 Den iboende kompleksiteten i minimalt liv: Å avdekke oddsen {kursiv og understreking ved oversetter.}
Den iboende kompleksiteten i minimalt liv: Å avdekke oddsen {kursiv og understreking ved oversetter.}
Oversatt fra Otangelo Grasso -Bilde 1; via FB, opprinnelig herfra
Livet på cellenivå viser komplekse molekylære prosesser, med hver komponent som spiller en viktig rolle i å opprettholde den livssentielle homeostasen og funksjonene til cellen. Disse prosessene, alt fra proteinsyntese til metabolisme, krever den sømløse koordinasjonen fra mange komplekse biomolekylære maskiner. Sentrale cellulære prosesser, som transkripsjon og oversettelse, står som testamenter for denne kompleksiteten. Disse systemene, interavhengige, fungerer gjennom presise molekylære språk og kodifisert, instruksjonsinformasjon og metainformasjon som inneholder kunnskapen, av når, og hvordan du kan utvinne og uttrykke den informasjonen for å lede fremstilling av alle viktige deler, som proteiner fra cellen. For eksempel signaliserer distinkte DNA -sekvenser oppstart og avslutning av transkripsjon. Videre oppnås cellulær koordinering gjennom omfattende signalveier som gjør det mulig for cellen å tilpasse seg og svare. Disse traséene er på sin side et komplekst nett av gjensidig avhengige komponenter. Reguleringsmekanismer orkestrerer ytterligere disse prosessene ved å avkode genetisk informasjon, kontrollere genuttrykk og sikre at cellulære prosesser ikke løper løpsk. Pelagibacter Ubque, de minste frittlevende bakteriene som er kjent i dag, opererer med omtrent 1.360 proteiner.
Bilde 2. Minimal celle med høye odds
 Hvert av disse proteinene er en sofistikert molekylær maskin med en spesifikk, livs-essensiell funksjon. Proteiner fungerer ofte bare hvis de er en del av komplekse produksjonskjeder som fungerer synergistisk. Oddsen for å danne en minimal celle av ren tilfeldighet er astronomisk lav: 1 til 10^530.280. Tatt i betraktning begrensningene i vårt univers, anses sannsynligheter utover 1 til 10^139 som statistisk umulige. Gitt denne rammen, er oddsen for en funksjonell celle som dannes tilfeldig ikke bare usannsynlig, men statistisk umulig. Dette bringer oss til en overbevisende konklusjon: den intrikate og gjensidig avhengige naturen til cellulære livspunkter, mer overbevisende i retning forsettlig design enn mot ikke-styrte hendelser.
Hvert av disse proteinene er en sofistikert molekylær maskin med en spesifikk, livs-essensiell funksjon. Proteiner fungerer ofte bare hvis de er en del av komplekse produksjonskjeder som fungerer synergistisk. Oddsen for å danne en minimal celle av ren tilfeldighet er astronomisk lav: 1 til 10^530.280. Tatt i betraktning begrensningene i vårt univers, anses sannsynligheter utover 1 til 10^139 som statistisk umulige. Gitt denne rammen, er oddsen for en funksjonell celle som dannes tilfeldig ikke bare usannsynlig, men statistisk umulig. Dette bringer oss til en overbevisende konklusjon: den intrikate og gjensidig avhengige naturen til cellulære livspunkter, mer overbevisende i retning forsettlig design enn mot ikke-styrte hendelser.
Det vitenskapelige samfunnet har historisk fremhevet enklere organismer som Mycoplasma for å understreke minimalistisk liv (som vil samsvare med et naturalistisk opprinnelse i livet), men dette perspektivet overser den iboende minimale kompleksiteten for livet til å starte. Mycoplasma, ofte utpekt som den minste livsformen, er symbiotisk og er avhengig av verten for å overleve og er derfor ikke en god kandidat og representant. En mer passende representasjon av minimalt frittlevende liv kan være bakterien, Pelagibacter Ubque, som kan skilte med omtrent 1.350 proteiner. Selv om 1.350 kan virke bare som et rent tall, er det avgjørende å gjenkjenne hvert protein som en kompleks molekylær maskin med livs-essensielle spesifikke funksjoner, som hver har en unik rolle. Mange av disse proteinene er intrikat koblet, og danner forseggjorte mikroproduksjonslinjer. Disse systemene fungerer ofte i harmoni, og utnytter synergi for å produsere viktige cellulære komponenter.
I min dyptgående utforskning av de grunnleggende behovene til de minste kjente livsformene, har jeg katalogisert en mengde proteiner og deres funksjoner for en antatt frittlevende celle i en hydrotermisk sjakt, som i dag er den dominerende hypotesen om hvor livet startet. Denne bestrebelsen alene har okkupert rundt 150 A4 sider -lenke.
Denne øvelsen gir et hint i størrelsen på intrikathet som ligger i selv de enkleste organismer.
Bilde 3. Problemer ved Livets Opprinnelse -oppsummert (O. Grasso)
 Nyere vitenskapspapirer, som Bill Martins fra 2021, poserer et minimalt metabolom bestående av 407 noder/proteiner, mens beregningene mine dobler det. I følge beregningene mine vil det være minst 862 noder/enzymer.
Nyere vitenskapspapirer, som Bill Martins fra 2021, poserer et minimalt metabolom bestående av 407 noder/proteiner, mens beregningene mine dobler det. I følge beregningene mine vil det være minst 862 noder/enzymer.
Hvis vi vurderer, at Lucas proteom og interaktom var et prøve- og feile -konsept, (som er alternativet til design) ville oddsen for å få en minimal størrelse med 1226 proteiner være:
Beregning av oddsen for å danne et proteom med 1 226 proteiner, hver av en gjennomsnittlig størrelse på 300 aminosyrer, ved rent tilfeldig prøving og feiling, er en monumental utfordring på grunn av den enorme antallet som er involvert. Her er en grunnleggende måte å tenke på oddsen på i en enkelt proteindannelse: Hver av de 20 vanlige aminosyrene har en 1 til 20 sjanse for å bli valgt til hver posisjon i proteinkjeden. For et protein med en sekvens på 300 aminosyrer, er sannsynligheten for å få en spesifikk sekvens ved en ren tilfeldighet: (1/20) ^300, eller 1 til 10 ^390. Hvis vi nå vurderer at vi trenger 1.360 slike spesifikke sekvenser, blir oddsen enda mer minuscule: ((1/20)^300)^1.360, eller 1 til 10^530.280.
Forutsetning 1: For at en funksjonell celle skal eksistere, krever det et intrikat og gjensidig avhengig system med koder, språk og proteiner, med oddsen for at disse systemene dannes tilfeldig 1 til 10^477.660.
Forutsetning 2: Sannsynligheter utover 1 av 10^139 (maksimalt antall mulige hendelser i et univers som er 13,8 milliarder år gammel (10^16 sekunder) der hvert atom (10^80) endrer tilstanden til maksimal hastighet av 10^40 ganger per sekund er 10^139.
KONKLUSJON: Derfor er den tilfeldige, ikke-styrte dannelsen av en funksjonell celles innbyrdes avhengige system i vårt univers statistisk umulig, noe som antyder forsettlig design.
Funksjonelle proteiner: Selv blant det enorme antallet mulige sekvenser vil bare en ekstremt liten brøkdel folde seg til funksjonelle proteiner. Så oddsen er enda verre når du vurderer dannelsen av funksjonelle proteiner. Mer om protein-dannelse her.
Bilde 4. Interaktom interaktiv samband innen en fungerende celle
 Interaksjoner og avhengigheter: Utover individuelle proteiner, må mange proteiner være i spesifikke former og krever spesifikke partnere eller kofaktorer for å fungere. Så kompleksiteten kommer ikke bare fra dannelsen av proteinene selv, men fra deres interaksjoner og avhengigheter.
Interaksjoner og avhengigheter: Utover individuelle proteiner, må mange proteiner være i spesifikke former og krever spesifikke partnere eller kofaktorer for å fungere. Så kompleksiteten kommer ikke bare fra dannelsen av proteinene selv, men fra deres interaksjoner og avhengigheter.
Samtidig forekomst: For en funksjonell celle vil mange av disse proteinene måtte komme til å eksistere samtidig, eller innen en levetid som gir mulighet for meningsfull interaksjon.
Den rene kompleksiteten til en enkelt celle og de astronomiske oddsene mot komponentene som kommer sammen ved ren tilfeldighet, viser frem den utrolige intrikatet og raffinementet som er innebygd i livet. Når vi dykker dypt inn i cellulære prosesser, blir den statistiske usannsynligheten ved at livet skulle opprinne og opererer gjennom ikke-styrte prosesser, ekstremt tydelig.
Ta oddsen for å danne et proteom med 1.360 proteiner, hver av en gjennomsnittlig størrelse på 300 aminosyrer, ved en ren tilfeldighet: 1 til 10^530.280. Dette tallet er uten tvil enormt. For å sette det i perspektiv, anslås det totale antallet atomer i det observerbare universet å være rundt 10^80. Bare for å illustrere størrelsen på dette tallet:
Hvis du stokker et standard kortstokk på 52 kort, er det omtrent 10^67 mulige arrangementer. Dette betyr at oddsen for å oppnå et spesifikt arrangement av proteiner er langt større enn oddsen for å blande et kortstokk til en spesifikk rekkefølge, ikke bare én gang, men milliarder og milliarder ganger etter hverandre.
Hvis hvert atom i det observerbare universet var et stoppeklokke, og hver stoppeklokke kunne telle til en billion (10^12) på bare ett sekund, og de alle teller samtidig fra begynnelsen av universet (for omtrent 14 milliarder år siden) til nå: ville vi fremdeles ikke ha kommet til 10^477.660. Faktisk ville vi ikke engang være i nærheten.
Oddsen for å danne disse proteinene etter tilfeldige prosesser er så hinsides at de er utenfor enhver tenkelig hendelse vi kan komme på i vårt univers. Dessuten stopper ikke kompleksiteten med bare å danne proteiner. Mange proteiner krever spesifikke former og konfigurasjoner, ellera vil de ikke fungere. Disse funksjonelle formene representerer en ekstremt liten undergruppe av alle mulige proteinformer. Derfor blir de allerede astronomiske oddsene for tilfeldig å generere enhver proteinsekvens enda mer usannsynlig, når de bare vurderer de sekvensene som resulterer i funksjonelle proteiner. Videre fungerer ikke proteiner isolert. De er en del av et intrikat nettverk av interaksjoner og avhengigheter. Mange proteiner krever spesifikke partnere, kofaktorer eller forhold for å utføre oppgavene sine. Dette betyr at flere proteiner samtidig må eksistere og samhandle på presise måter for cellulære funksjoner å fortsette. Produksjons-, signal- og reguleringskoder og språk som ligger til grunn for cellulære prosesser eksemplifiserer ikke-reduserbarhet og gjensidig avhengighet. Tenk på den genetiske koden, der DNA -sekvenser blir transkribert til RNA og deretter oversatt til proteiner. Hvert av disse trinnene er avhengig av en pakke med molekylære maskiner, og hver del av dette maskineriet er viktig. Uten transkripsjon forblir informasjonen i DNA låst. Uten oversettelse er meldingene i RNA meningsløse. Disse prosessene er avhengige av hverandre, og den ene er meningsløs uten den andre. Videre tillater språkene og kodene til cellen intrikat kommunikasjon og kryss-kommunikasjon. Signalveier lar celler svare miljøet, regulere genuttrykk og koordinere aktiviteter. Uten disse kommunikasjons-nettverkene ville cellen være en samling av deler uten koordinering.
Det cellulære maskineriet trenger også reguleringsmekanismer. Uten regulering kan cellulære prosesser gå amok, noe som fører til sykdom eller celledød. Reguleringsmekanismene sikrer at alt skjer når det skal, hvor det skal og i riktige mengder. Det er utfordrende å se for seg hvordan slike gjensidig avhengige systemer kunne ha dukket opp trinn for trinn over millioner, til og med milliarder, av år. Hvis én del av systemet manglet eller ikke er fullt funksjonelt, vil hele systemet sannsynligvis mislykkes. Disse systemene er krystallklare bevis på at alle komponenter som kreves for å være til stede fra begynnelsen, noe som antyder at de var designet for å fungere som sammenkoblede helheter.
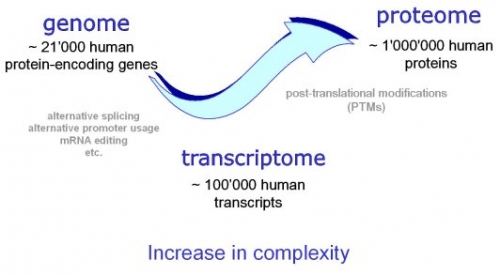
Bilde 5. Proteom
 Luca er en teoretisk enhet, en encelleorganisme som angivelig alle livsformer på jorden stammet fra gjennom evolusjon.
Luca er en teoretisk enhet, en encelleorganisme som angivelig alle livsformer på jorden stammet fra gjennom evolusjon.
Et minimalt proteom for den siste universelle felles stamfaren (LUCA): Følgende sekvens starter med grunnleggende biokjemiske reaksjoner og cellulær energiproduksjon, og går deretter videre til informasjonsbærere, strukturelle komponenter og til slutt forsvars- og reparasjonsmekanismer.
Metallklynger: Enzymer/proteiner Estimat: 46 -Viktig for forskjellige biokjemiske reaksjoner og proteinstrukturer.
Energimetabolisme, sentral karbonmetabolisme og andre spesifikke veier: enzymer/proteiner estimat: 74 -Grunnleggende stier som gir energi og forløpere for andre biosyntetiske prosesser:
Nukleotidsyntese og reparasjon: enzymer/proteiner estimat: 89 -Grunnlaget for generering av genetiske informasjonsbærere:
Aminosyre-biosyntese: Enzymer/proteiner estimat: 135 -Byggesteiner for proteinsyntese:
Reguleringsenzymer og proteiner i aminosyresyntese: enzymer/proteiner estimat: 76 -Regulering av aminosyre-syntese.
Oversettelse/ribosom i Luca: Enzymer/proteiner Estimat: 125 -Prosesser og maskiner for proteinsyntese.
Biosyntese og montering av bakteriell ribosom: Enzymer/proteiner Estimat: 104 -Ytterligere utdyping av ribosommontering og funksjon.
Transkripsjon/regulering i LUCA: Enzymer/proteiner Estimat: 63 -Prosesser for å lese genetisk informasjon og regulering.
DNA -prosessering i Luca: Enzymer/proteiner estimat: 48 -Administrere og replikere genetisk informasjon.
Familier/funksjoner involvert i forskjellige aspekter ved celledeling i LUCA:Enzymer/proteiner estimat: 96 - Celledeling og formering:
Peptidoglycan syntese: Enzymer/proteiner estimat: 91 -Essensielt for syntese av bakteriell cellevegg.
Fettsyre og fosfolipidsyntese i Luca: Enzymer/proteiner estimat: 48 -For å lage cellemembraner.
Kofaktorer: Enzymer/proteiner Estimat: 85 -Viktige hjelpere for enzymatiske reaksjoner.
NAD metabolisme: Enzymer/proteiner estimat: 63 -Viktig for redoksreaksjoner i cellen.
Reaktive OkSygenarter (ROS): Enzymer/proteiner estimat: 3 -Håndtere oksidativt stress og biprodukter av metabolisme.
Ukarakterisert: 136.
Den totale summen for alle oppgavene som er gitt er: 1.360.
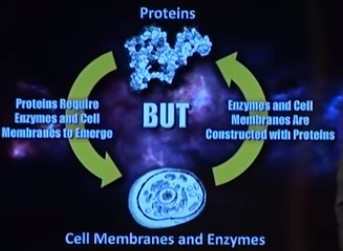
Bilde 6. Høne-egg problem Prtoeiner vs enzymer og celle-membraner
 Antallet 1 226 proteiner er i størrelsesordenen til hva som kan betraktes som et minimalt proteom. For kontekst er bakterien Pelagibacter Ubque (medlem av SAR11 -kleden, en av de mest tallrike og minste marine mikrober). P. Ubque har også et lite genom (1 308 759 bp), for tiden det minste genomet som er kjent for en frittlevende organisme. Genomet koder for 1 354 forutsagte proteiner, 1 rRNA -operon og 32 tRNA. Gitt at P. ubique har et strømlinjeformet genom tilpasset for sitt spesifikke oseaniske miljø, er proteintallet nær den nedre grensen for frittlevende organismer.
Antallet 1 226 proteiner er i størrelsesordenen til hva som kan betraktes som et minimalt proteom. For kontekst er bakterien Pelagibacter Ubque (medlem av SAR11 -kleden, en av de mest tallrike og minste marine mikrober). P. Ubque har også et lite genom (1 308 759 bp), for tiden det minste genomet som er kjent for en frittlevende organisme. Genomet koder for 1 354 forutsagte proteiner, 1 rRNA -operon og 32 tRNA. Gitt at P. ubique har et strømlinjeformet genom tilpasset for sitt spesifikke oseaniske miljø, er proteintallet nær den nedre grensen for frittlevende organismer.
Noen generelle kategorier og enzymer som kan anses som grunnleggende for et bredt spekter av cellulære livsformer:
Membrantransportsystemer: Mens noen transportører ble nevnt, ville en organisme trenge transportsystemer for ioner, vann (aquaporins) og andre essensielle molekyler som ikke er spesifisert i listen din.
ATP -syntese: ATP -syntasekomplekset, avgjørende for å produsere ATP, den primære energivalutaen til cellen, ble ikke nevnt.
DNA -replikasjonsmaskiner: De grunnleggende DNA -polymerase -enzymer, helikaser, primaser, ligaser og topoisomeraser, som er involvert i DNA -replikasjon og vedlikehold, var ikke detaljert.
Proteinfolding og nedbrytning: chaperones (som Groel/Groes og DNAK/DNAJ) hjelper til med proteinfolding. Proteasom- eller CLPXP -maskiner er med på å degradere unødvendige eller feilfoldede proteiner.
Glykolyse og TCA -syklus enzymer: sentrale karbonmetabolisme -enzymer, som de i glykolyse og TCA (Krebs) -syklusen -lenke, gir viktige forløpere for forskjellige biosyntetiske veier og produserer ATP.
Celledelingsmaskiner: FTSZ og andre proteiner som er essensielle for celledeling og septumdannelse.
Stressresponssystemer: Ulike enzymer og proteiner hjelper celler med å takle oksidativt stress, DNA -skade og andre miljøs stressfaktorer. For eksempel hjelper SuperOksyd Dismutase (SOD) og katalase med å nøytralisere reaktive oksygenarter.
Lipopolysakkaridsyntese: I gramnegative bakterier er enzymer involvert i lipopolysakkaridsyntese viktige for ytre membran-biogenese.
RNA -prosessering og nedbrytning: Mens du nevnte ribonukleaser, kan andre enzymer og proteiner assosiert med RNA -spleising, modning og nedbrytning i forskjellige organismer inkluderes.
Signaltransduksjonssystemer: To-komponentsystemer, proteinkinaser og andre signalmolekyler hjelper cellen til å svare på miljøendringer.
Autotrofiske prosesser: Hvis du vurderer autotrofer, kan Calvin -syklusen -lenke og andre karbonfikseringsstier, sammen med enzymer for nitrogenfiksering, anses som essensielle.
Bilde 7. Metabolomet -plass og funksjon -By Jmarchn - Own work, CC BY-SA 3.0
 Størrelse på det sentrale metabolomet
Størrelse på det sentrale metabolomet
Det sentrale metabolomet refererer generelt til et sett med metabolske veier og prosesser som er allestedsnærværende og grunnleggende for cellulær liv. Basert på den forståelsen, kan følgende betraktes som en del av det sentrale metabolomet:
Energimetabolisme, sentral karbonmetabolisme og andre spesifikke veier: enzymer/proteiner estimat: 74 -Grunnleggende veier som gir energi og forløpere for andre biosyntetiske prosesser.
Nukleotidsyntese og reparasjon: enzymer/proteiner estimat: 89 -Grunnlaget for generering av genetiske informasjonsbærere.
Aminosyre-biosyntese: Enzymer/proteiner estimat: 135 -Byggesteiner for proteinsyntese.
Reguleringsenzymer og proteiner i aminosyresyntese: enzymer/proteiner estimat: 76 -Regulering er avgjørende for å opprettholde metabolsk homeostase og sikre effektiv bruk av cellulære ressurser.
Oversettelse/ribosom i Luca: Enzymer/proteiner Estimat: 125 -Prosesser og maskiner for proteinsyntese.
Biosyntese og montering av bakterielt ribosom: Enzymer/proteiner Estimat: 104 - Ytterligere utdyping av ribosommontering og funksjon.
Transkripsjon/regulering i LUCA: Enzymer/proteiner Estimat: 63 -Prosesser for å lese genetisk informasjon og regulering.
DNA -prosessering i Luca: Enzymer/proteiner estimat: 48 -Administrere og replikere genetisk informasjon.
Kofaktorer: Enzymer/proteiner Estimat: 85 -Viktige hjelpere for enzymatiske reaksjoner.
NAD metabolisme: Enzymer/proteiner estimat: 63 -Viktig for redoksreaksjoner i cellen.
Totalt 862 enzymer/proteiner
Bilde 8. Utfordringer ved Opprinnelse Til Liv (OTL-abiogenese)
 De andre prosessene og enhetene som er oppført er viktige for livet, men de kan betraktes som mer spesialiserte, snarere enn en del av den sentrale kjernen i metabolismen. For eksempel, mens "metallklynger" og "Reaktive OkSygenarter (ROS)" er avgjørende for mange organismer, kan de bli sett på som tillegg til de sentrale metabolske prosessene i noen sammenhenger.
De andre prosessene og enhetene som er oppført er viktige for livet, men de kan betraktes som mer spesialiserte, snarere enn en del av den sentrale kjernen i metabolismen. For eksempel, mens "metallklynger" og "Reaktive OkSygenarter (ROS)" er avgjørende for mange organismer, kan de bli sett på som tillegg til de sentrale metabolske prosessene i noen sammenhenger.
Ytterligere betraktninger
Den intrikate dansen av molekylære prosesser i en celle viser et orkester av sammenvevdd systemer, språk og koder. Den rene kompleksiteten og presisjonen til disse prosessene antyder et grunnleggende mysterium: Hvordan kunne slik intrikathet oppstå trinn for trinn, når mellomtrinnene ikke ville ha noen funksjon? Ta for eksempel prosessen med transkripsjon og oversettelse. Transkripsjon leser genetisk informasjon, og translasjon konverterer den meldingen til proteiner. Hvert av disse trinnene er avhengig av en serie med molekylær maskiner. Uten transkripsjon forblir informasjonen i DNA utilgjengelig. Uten oversettelse tjener meldingene i RNA ingen formål. De to prosessene er avhengige av hverandre, og gjør den ene meningsløs uten den andre. Videre er språkene og kodene som understøtter disse prosessene ikke-reduserbare i deres kompleksitet. Den genetiske koden, med sine spesifikke sett med bokstaver, ord og regler, må synkroniseres perfekt med translasjonsmaskineriet. En endring eller fravær i én, ville gjøre de andre nonsensiske. Så er det cellulær signalering, et stort nettverk av kommunikasjonskanaler som koordinerer enhver cellulær aktivitet. Signalveier gjør det mulig for celler å føle miljøet, regulere genuttrykk og orkestrere komplekse oppgaver. Uten disse traséene ville en celle bare være et utvalg av deler uten direktiv. Videre kan disse signalnettverkene ikke fungere isolert. De krever spesifikke reseptorer, sekundære budbringere og effektorproteiner - som alle er avhengige av hverandre. Fravær eller funksjonsfeil i en komponent kan gi følgefeil gjennom systemet og forårsake utbredt dysfunksjon. Men kompleksiteten stopper ikke der.
Bilde 9. Otangelo konkluderer om livets opprinnelse

Reguleringsmekanismer styrer disse systemene, og sikrer at alt oppstår når og hvor det skal. Denne intrikate koreografien krever et sofistikert sett med koder og språk. For eksempel en bestemt sekvens av DNA, kjent som en promoter, signaler der transkripsjonen skal begynne. En annen sekvens kalt en terminator, signaliserer der den skal ende. Disse reguleringskodene er ikke-reduserbare; Uten dem ville transkripsjon enten ikke starte, eller produsere meningsløse sekvenser. Det er utfordrende å forestille seg hvordan slike sammenflettede systemer kunne ha oppstått gjennom en lineær, trinnvis prosess, selv over milliarder av år. Mellomstadier, som mangler full funksjonalitet, ville ikke gi en overlevelsesfordel. Den gjensidige avhengigheten av disse systemene antyder at de måtte være til stede og fullt funksjonelle fra starten. Når vi dykker dypt inn i disse systemene, blir det tydelig at de ikke kunne ha dukket opp trinn for trinn. Den ene uten den andre er som en lås uten nøkkel - ubrukelig. Denne sammenkoblede kompleksiteten peker på et design som er bevisst og forsettlig, og krever en dyp forståelse av cellulære prosesser. Avslutningsvis er den molekylære verden i en celle ikke bare et virvar av reaksjoner. Det er en harmonisert symfoni av koder, språk og systemer som er ikke-reduserbare, kompliserte og gjensidige avhengige. Brukervennligheten av en slik koordinert dans som dukker opp gjennom tilfeldige, trinnvise prosesser, understreker vidunderet av cellulært liv og antyder en intelligent orkestrering bak dets eksistens.
Oversettelse via google oversetter og bilder ved Asbjørn E. Lund